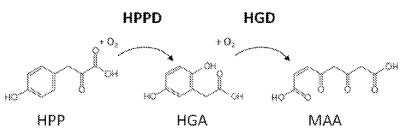
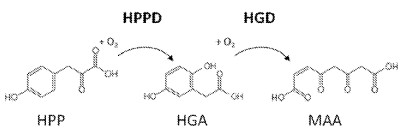
„4-Hydroxyphenylpyruvate dioxygenase (HPPD, EC 1.13.11.27) is an enzyme engaged in photosynthetic activity and catalyzes the transformation of 4-hydroxyphenylpyruvic acid (HPPA) into homogentisic acid (HGA). HPPD inhibitors constitute a promising area of discovery and development of innovative herbicides with some advantages, including excellent crop selectivity, low application rates, and broad-spectrum weed control. HPPD inhibitors have been investigated for agrochemical interests, and some of them have already been commercialized as herbicides.“
[0004] Inhibition of HPPD leads to uncoupling of photosynthesis, deficiency in accessory light-harvesting pigments and, most importantly, to destruction of chlorophyll by UV-radiation and reactive oxygen species (bleaching) due to the lack of photo-protection normally provided by carotenoids (Norris et al. (1995), Plant Cell 7: 2139-2149). Bleaching of photosynthetically active tissues leads to growth inhibition and plant death.
[0005] Some molecules which inhibit HPPD (hereinafter named HPPD inhibitor herbicides), and which inhibit transformation of the HPP into HGA while binding specifically to the enzyme, have proven to be very effective herbicides.
[0006] At present, most commercially available HPPD inhibitor herbicides belong to one of these chemical families, as listed below:
1) the triketones, e.g. …; 2) the diketonitriles ..; 3) the isoxazoles ..; 4) the hydroxypyrazoles; ..; ..
[0025] In the present invention, HPPD polypeptides and plants containing them, showing a full tolerance against one or more HPPD inhibitor herbicides belonging to various chemical classes, are described. It turned out that in order to generate such HPPD polypeptides with maximized and broad tolerance against several classes of HPPD inhibitor herbicides, it is important to reduce the affinity to the HPPD polypeptide (ki) concerning the respective HPPD inhibitor herbicide(s) and simultaneously to ensure an improved rate of dissociation (koff) of a slow-binding or slow, tight-binding inhibitor as known from the wild-type and several mutant HPPD polypeptides to achieve high level of inhibitor tolerance.
In the present invention, this goal was achieved by developing a set of HPPD polypeptides, which have either no or only a significantly reduced affinity to HPPD inhibitor herbicides and, at the same time, the rate of dissociation of the HPPD inhibitor herbicides of the enzyme is increased to such an extent that the HPPD inhibitor herbicides no longer act as slow-binding or slow, tight-binding inhibitors but, instead of this, have become fully reversible inhibitors. In the present invention, compositions and methods for obtaining a new set of HPPD polypeptides having the before mentioned characteristics (i.e. no or only a significantly reduced affinity to HPPD inhibitor herbicides, increased rate of dissociation of the HPPD inhibitor herbicides of the enzyme; HPPD inhibitor herbicides no longer act as slow-binding or slow, tight-binding inhibitors but have become fully reversible inhibitors) are provided. Compositions include HPPD polypeptides and isolated, recombinant or chimeric nucleic acid molecules encoding such HPPD polypeptides, vectors and host cells comprising those nucleic acid molecules. Compositions also include the antibodies to those polypeptides. The nucleotide sequences can be used in DNA constructs or expression cassettes for transformation and expression in organisms, including microorganisms and plants. The nucleotide sequences may be synthetic sequences that have been designed for expression in an organism including, but not limited to, a microorganism or a plant.
[0026] The compositions include nucleic acid molecules encoding herbicide tolerant HPPD polypeptides, including nucleic acid molecules encoding an HPPD polypeptide having (a) a proline at the amino acid position corresponding to amino acid position 335 of SEQ ID NO:1, (b) a histidine or an aspartic acid at the position corresponding to amino acid position 336 of SEQ ID NO:1, and (c) a serine at the position corresponding to amino acid position 337 of SEQ ID NO:1 and, optionally, one or more further amino acid substitutions at the positions corresponding to amino acid positions 204, 213, 264, 268, 270, 310, 315, 330, 331, 338, 339, 340, 344, 345 of SEQ ID NO: 1, including the HPPD polypeptides set forth in any of SEQ ID NO:3-108 as well as fragments thereof.
[0029] FIG. 1 shows a simplistic scheme of the coupled HPPD activity assay used in this invention to determine the enzymatic activity of the exemplary HPPD polypeptides.
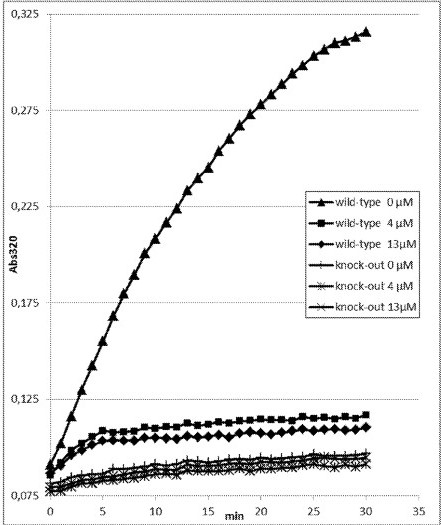
[0030] FIG. 2 shows exemplary kinetic changes in absorbance at 320 nm (Abs320) in raw extracts samples of wild-type and knock-out HPPD polypetide observed with 200 μM HPP and 0, 4 or 13 μM Cmpd. 1 (2-chloro-3-(methylsulfanyl)-N-(1-methyl-1H-tetrazol-5-yl)-4-(trifluoromethyl)benzamide) according to Example 3 in the coupled HPPD activity assay. The knock-out HPPD polypeptide was obtained by exchanging a histidine to an alanine at the amino acid position corresponding to amino acid position 162 of SEQ ID NO:1. This position is well known for its importance due to its involvement in the coordinated binding of the iron atom in the active site of the HPPD polypeptide (Serre et al. (1999), Structure, 7, 977-988).
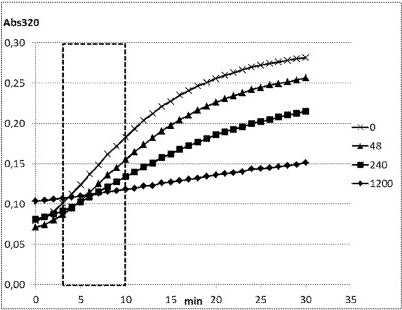
[0031] FIG. 3 shows exemplary kinetic changes in absorbance at 320 nm (Abs320) of a purified mutant HPPD polypeptide corresponding to SEQ ID NO:17 according to Example 3 observed at high substrate concentration and with 0, 48, 240 or 1200 μM Cmpd. 2 (2-methyl-N-(5-methyl-1,3,4-oxadiazol-2-yl)-3-(methylsulfonyl)-4-(trifluoromethyl)benzamide) in the coupled HPPD activity assay. The apparent kinetic constant (kapp) was determined as signal change over time (delta_Abs320/min) in the boxed timeframe.
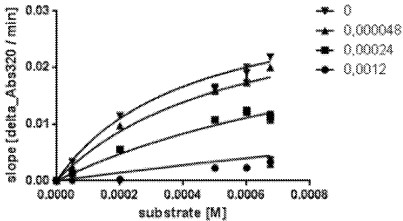
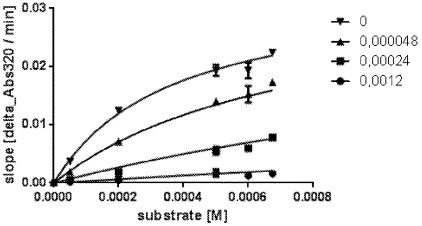
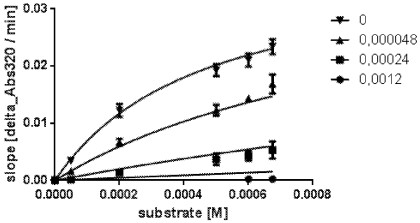
[0032] FIG. 4 depicts data from an exemplary ki determination with a purified mutant HPPD polypeptide corresponding to SEQ ID NO:17 with different inhibitor and substrate (HPP) concentrations by fitting according to the competitive inhibition model:
[0033] a) Kinetic changes in absorbance at 320 nm over time (delta_Abs320/min) in the presence of 0-0.0012 M of Cmpd. 2 at the given substrate concentration, according to Example 3;
[0034] b) .. of Cmpd. 1 (2-chloro-3-(methylsulfanyl)-N-(1-methyl-1H-tetrazol-5-yl)-4-(trifluoromethyl) benzamide) ..
[0035] c) .. of MST ..
[0036] d) .. of DKN …
[0037] All exemplary HPPD polypeptides, which are summarized in Tables 2, 3, 4, and 5 were measured and analyzed as shown for example with SEQ ID NO:17 in FIGS. 3 & 4.
Example 1. Creation of Mutated HPPD Polypeptides by Site-Directed Mutagenesis
[0240] The Pseudomonas fluorescens HPPD nucleotide sequence (SEQ ID NO:109) as described in WO2009/144079 encoding the HPPD polypeptide corresponding to SEQ ID NO:1 was cloned according to well known methods in the art and as described in WO2014/043435. Subsequent site-saturated mutagenesis, site-directed mutagenesis and combinatorial variants with one or more mutations of the nucleic acid encoding sequence of wild-type PfHPPD polypeptide encoding the recombinant HPPD polypeptide corresponding to SEQ ID NO: 1 were carried out using standard PCR-based technologies well known in the art (and as described likewise in WO2014/043435). All designed and synthesized mutant clones were confirmed by DNA sequencing using plasmid specific oligonucleotides. Table 2, below, summarizes the exemplary mutant HPPD polypeptides (SEQ ID NO:2-SEQ ID NO:108).
{Es folgt eine Tabelle mit Aminosäureaustauschen in 108 HPPD-Polypeptiden}
Example 2: Cloning, Expression, and Purification of Recombinant Wild-Type and Mutant HPPD Polypeptides
[0242] All resulting nucleic acid encoding sequences of wild-type and mutant HPPD encoding the recombinant HPPD polypeptide were cloned, produced and purified using methods well known in the art (Sambrook et al., Molecular Cloning: A Laboratory Manual, 3rd ed., CSH Laboratory Press, 2001, Cold Spring Harbor, N.Y.). All resulting nucleic acid encoding sequences were cloned into pSE420(RI)NX fused with an N-terminal His-tag (encoding the amino acid sequence M1-A2-H3-H4-H5-H6-H7-H8-), as described in WO2014/043435, and were expressed in Escherichia coli strain BL21 (DE3) (New England Biolabs, Frankfurt, Germany). For clarity, all listed positions with the respective amino acid exchanges from mutant HPPD polypeptides in Tables 1 to 5, and Table 7 corresponding to SEQ ID NO:2 to SEQ ID NO:108 in this invention, refer to the native wild-type HPPD amino acid sequence without the N-terminal His-tag corresponding to SEQ ID NO:1.
[0243] For the generation of purified HPPD polypeptide samples, cells were grown for 3 h at 37° C. in 5 ml LB medium containing 100 μg/ml ampicillin in a 50 ml shaker flask at 140 rpm. One ml of this starter culture was used as inoculum for the expression culture. Cells were grown for about 3 h at 37° C. in 100 ml LB medium containing 100 μg/ml ampicillin and 150 mM Hepes (Merck, Darmstadt, Germany) in a 500 ml shaker flask at 120 rpm. At an OD600 of about 0.6, IPTG (Roth, Karlsruhe, Germany) was added to a concentration of 0.4 mM. After further growth for 60 min at 37° C., the temperature was reduced to 28° C. and growth continued for another 18 h at 140 rpm. Cells were harvested by centrifugation at 4° C., 3200 g for 30 min in 50 ml Falcon tubes and cell pellets were stored at −80° C. Cells were lysed and his-tagged protein was purified according to manufacturer protocol of the used Ni-NTA Fast Start Kit (Qiagen, Hilden, Germany) with following adaptions for increased yield: cells from 50 ml culture were lysed in 4 ml and lysate supernatant was generated by centrifugation for 15 min at 18000 g. The amount of matrix in the columns was increased by addition of 1 ml of NiNTA Superflow (Qiagen, Hilden, Germany) each and extensively re-buffered into 20 mM Tris (pH 7.6) (Merck, Darmstadt, Germany) Lysate supernatant was applied and His-tagged protein was bound to the Ni-NTA matrix by incubation for 1 h at 4° C.
[0244] The resulting protein samples were re-buffered into 20 mM Tris, 20% Glycerol (pH 7.8) (Sigma-Aldrich, St. Louis, USA) by use of Zeba™ Spin Desalting Columns, 7K MWCO, 10 mL (Thermo Fisher Scientific, Waltham, USA) and analysed for protein concentration and purity by Abs280 (NANODROP 8000, Thermo Fisher Scientific, Waltham, USA) and SDS-PAGE. The concentrations of purified proteins were generally in the range of 0.6-4.6 mg/ml by an estimated purity of about 90%.
[0245] For the generation of crude HPPD polypeptide extract in micro titer plates (MTP) for the determination of residual activity in inhibition assays, cells were grown in 40 or 150 μl LB medium containing 1% Glucose (Merck, Darmstadt, Germany) and 100 μg/ml ampicillin in a standard 96 well plate (Thermo Fisher Scientific, Waltham, USA) incubated for about 18 h in a humidity incubator at 37° C.
[0246] 30 μl of this starter culture were added to 600 μl LB medium containing 100 μg/ml ampicillin and 150 mM Hepes (Merck, Darmstadt, Germany) as inoculum for the expression culture in 96 well plates (2 ml deep wells; HJ Bioanalytik, Erkelenz, Germany) The plates were sealed by an aluminium foil, and cells were incubated for 5 h at 37° C. on a plate shaker at 750 rpm. The expression was induced by addition of IPTG in a final concentration of 1 mM followed by further sealed incubation for about 18 h at 30° C. on a plate shaker at 750 rpm.
[0247] Cells were harvested by centrifugation at 4° C., 2500 g for 15 min discarding the supernatant. Cell pellets were stored at −80° C. and lysed in 250 μl 1× BugBuster® (Merck, Darmstadt, Germany) in 140 mM Tris (pH 7.8), with 1:25000 diluted BNase® (Qiagen, Hilden, Germany) by incubation of the resuspended cells for 30 min at 4° C. and 1000 rpm. Lysates were clarified by centrifugation for 15 min at 4° C., 2500 g, and 150 μl supernatant were transferred in standard 96 well plate (Thermo Fisher Scientific, Waltham, USA) for subsequent testing in quadruplets.
Example 3: HPPD Enzyme Assay to Analyse Mutant HPPD Polypeptides with Improved HPPD Inhibitor Herbicide Tolerance
[0248] The activity of HPPD polypeptides was determined in absence or presence of HPPD inhibitors using the coupled HPPD activity assay (FIG. 1).
[0249] For the determination of the residual activity, the apparent kinetic constant (kapp) of the determined substrate conversion was measured as kinetic changes in absorbance at 320 nm in a coupled assay, in that homogentisate (HGA) formed by HPPD from HPP is directly converted into the well absorbing molecule maleylacetoacetate (MAA) by a second enzyme homogentisate dioxygenase (HGD), applied in excess uniformly in all assays (see FIG. 1). The measurements were performed in 384 micro titer plates (Greiner Bio-One GmbH, Frickenhausen, Germany) by plate readers (Tecan infinite M1000 or M1000PRO, Tecan, Männedorf, Switzerland). The kcat/kM ratio of an enzymatic activity is proportional to the apparent kinetic constant kapp and is proportional to kcat/kM *[E] ([E]=enzyme concentration).A competitive inhibitor exhibits an apparent increase in kM and thereby a reciprocal decrease in kapp at non-saturating substrate concentrations. As both kapp measurements in the presence and absence of inhibitor were performed by use of the identical enzyme sample, crude or purified, and thereby at the same enzyme concentration, the enzyme concentration eliminates from the calculation of residual activity and the ratio of both kapp directly indicates the change of kM due to the inhibition. Noteworthy, this concept applies to enzyme/inhibitor pairs interacting in a “competitive inhibition” manner, probably correct for almost all polypeptide variants and inhibitors described herein, but for sure not with respect to the wild-type polypeptide, which is inhibited irreversibly (for comparison see WO2014/043435, FIGS. 2 and 3;). Consequently, residual activities of the wild-type HPPD polypeptide referring to “competitive inhibition” and ki values can’t be correctly calculated, nevertheless, for the purpose of illustration unfounded values are given in Table 3 for the wild-type HPPD polypeptide, which are calculated by initial changes in signal before the irreversible inhibition took place.
[0250] The assay solution used for determination of residual activities in raw HPPD polypeptide samples was composed by 200 mM sodium phosphate (Merck, Darmstadt, Germany, pH 7.0), 5 mM MgCl2 (Merck, Darmstadt, Germany), 20 mM ascorbate (Sigma-Aldrich, St. Louis, USA), 1 mM DTT (Sigma-Aldrich, St. Louis, USA), 0.1% Pluronic F-68 (Sigma-Aldrich, St. Louis, USA), 40 μM FeSO4 (Sigma-Aldrich, St. Louis, USA), about 8 mg/ml purified HGD and low or high concentrations of substrate HPP (100 or 400 μM) from a 1 M stock solution in DMSO (Sigma-Aldrich, St. Louis, USA) and equilibrated for 20 min on ice. For every HPPD polypeptide sample two assays were performed in quadruplets, whereby 5 μl of HPPD polypeptide sample were mixed firstly with 5 μl buffer (1× BugBuster®; (Merck, Darmstadt, Germany); in 140 mM Tris, pH7.8, with 1:25000 diluted BNase®; Qiagen, Hilden, Germany)) or 5 μl inhibitor diluted in the same buffer from a 0.1 M stock solution in DMSO (30, 100 or 120 μM resulting in 15, 50 and 60 μM in the HPPD polypeptide/inhibitor sample) in the reference and inhibition assay, respectively, and subsequently with 10 μl assay solution. The change in absorbance at 320 nm was followed in 1 min intervals for 30 min. The kapp values were calculated as signal slope over time in the early phase of the kinetic reaction, usually for the first 5-10 minutes of the measurements (compare FIG. 3). Additionally and according to the calculated residual activity, the total conversion, i.e. the absolute change in signal, in the 30 min timeframe was monitored as measure of turnover, and a residual turnover was calculated by dividing the change in signal in the presence of inhibitor by the change in signal in the reference sample without inhibitor.
[0251] The assay solutions used for determination of ki values were composed in the same way containing six different concentrations of HPP substrate (0-1350 μM) for each of the four inhibitor concentrations tested. The inhibitors were diluted in 140 mM Tris, 0.05% Pluronic F-68 (Sigma-Aldrich, St. Louis, USA) and applied in concentrations adopted for the respective HPPD polypeptide/inhibitor pairs to generate dynamic data (FIG. 4a-d); generally, their concentrations in the HPPD polypeptide/inhibitor sample were in the range from 0 to 0.0012 M.
Example 4: Improved Herbicide Tolerance Mediated by Residue Exchanges in HPPD Polypeptides
[0252] When the tolerance of mutant HPPD polypeptides was determined against different available chemical classes of HPPD inhibitor herbicides (triketones, isoxazoles, N-(1,3,4-oxadiazol-2-yl)benzamides, or N-(tetrazol-5-yl)-arylcarboxamides), it became evident that some of the new embodiments in this invention are not only significantly improved compared to reference wild-type HPPD (SEQ ID NO:1), but also unexpectedly better than the prior art mutant HPPD polypeptides (like, for example, those being disclosed in WO2014/043435) with SEQ ID NO:2 in this invention as an example.
[0253] As outlined in Table 3, prior art mutant HPPD polypeptides (WO2014/043435) corresponding to SEQ ID NO:2 in this invention contains residue exchanges at position 335, 336, 339 and 340. Based on mutant HPPD polypeptide comprising 335 (E=>P), 336 (G=>D/H), 337 (N=>S), the introduction of further residue exchanges at position 264, 268, 270, 330, 340 and/or 345 generated mutant HPPD polypeptides showing strongly improved tolerance (Table 3), concerning multiple applied HPPD inhibitors belonging to various chemical classes.
{Tabelle 3 listet 31 HPPD-Polypeptide und ihre Herbizidtoleranz (3a-d): „Tolerance of mutant HPPD polypeptides against different HPPD inhibitor herbicides belonging to diverse chemical classes.“}
[0254] Accordingly, we generated and evaluated new mutant HPPD polypeptides by combinatorial residue exchanges at position 335 (glutamic acid=>proline), 336 (glycine=>aspartic acid/histidine), 337 (asparagine=>serine) and, optionally, further comprising exchanges at position 204, 213, 264, 268, 270, 310, 315, 330, 331, 338, 339, 340, 344 and/or 345 (Table 3, 4 and 5), that exhibit improved residual activities, higher residual turnover, higher ki values and thereby significantly higher herbicide tolerance. Depending on the HPPD inhibitor herbicide tested, the level of improvement might differ concerning the HPPD polypeptides employed in such assay, with a level of 1.5 up to 110 fold, compared to SEQ ID NO: 2 (Table 4).
[0255] Analysis of the time-course of inhibition against the different HPPD inhibitor herbicide chemical classes revealed, that the HPPD inhibitor herbicides appear to be reversible inhibitors against the new mutant HPPD polypeptides, in contrast to the slow and tight binding inhibitor characteristic of the wild-type HPPD polypeptide corresponding to SEQ ID NO:1 (see FIGS. 2 and 3). These behaviors provide a better and versatile potential for tolerances in crop plants to various HPPD inhibitor herbicides.
[0256] For high residual activity in the presence of HPPD inhibitor herbicides, the disclosed positions and residue changes are highlighted in Table 1 relative to the amino acid position in the HPPD polypeptide corresponding to SEQ ID NO:1 in this invention are shown to be important.
[0257] Residual activities and residual turnover were determined according to Example 3 by measuring kapp and total change in signal, respectively, in the presence and absence of (a) Cmpd. 1 (2-chloro-3-(methylsulfanyl)-N-(1-methyl-1H-tetrazol-5-yl)-4-(trifluoromethyl)benzamide), (b) Cmpd. 2 (2-methyl-N-(5-methyl-1,3,4-oxadiazol-2-yl)-3-(methylsulfonyl)-4-(trifluoromethyl)benzamide), (c) mesotrione (MST), and (d) diketonitrile (DKN). For each mutant HPPD polypeptide, the kapp and total change in signal without HPPD inhibitor herbicides served for normalization of the kapp and total change in signal in the presence of the herbicide. The summarized resulting “%-values” in the respective tables 3a), 3b), 3c), and 3d) are means of two independent experiments with an average standard deviation of 5%. The reaction was performed at high substrate concentrations with Cmpd. 1 and Cmpd. 2 at a concentration of 15 μM and the other two herbicides (DKN, MST) at a concentration of 60 μM. For clarity, empty cells at the respective amino acid position in SEQ ID NO:2 to SEQ ID NO:108 are defined as identical to the amino acids corresponding to SEQ ID NO:1, highlighting only the exchanges in the HPPD polypeptide variant. The abbreviation “n.i.” means that no inhibition was observed under the given conditions, i.e. the kapp or the total change in signal in the presence of inhibitor is not decreased compared to the corresponding value in the absence of inhibitors.
[0258] The mutant HPPD polypeptides corresponding to SEQ ID NO:10 and SEQ ID NO:24 with amino acid exchanges at positions 335, 336, 337, 340, and 345 relative to HPPD polypeptide according to SEQ ID NO:1, exhibit in the presence of various HPPD inhibitors tested a significant improvement regarding residual activities and residual turnovers (Table 3a-d). The depicted herbicide tolerance of SEQ ID NO:10 shows not only the improvement vs. the wild-type HPPD polypeptide (SEQ ID NO:1), but also vs. the prior art (WO2014/043435) mutant HPPD polypeptide corresponding to SEQ ID NO:2 in this invention across all four depicted different herbicide classes.
[0259] Further improvements in HPPD inhibitor tolerance are apparent in variants with residue exchanges at the disclosed amino acid positions 268 and 270.
[0260] Starting from SEQ ID NO: 24, SEQ ID NO:31 differ only at the stated positions 268 and 270. These changes increase significantly the residual turnover in the presence of HPPD inhibitor Cmpd. 1 and MST (Table 3a and 3c), and keeps the already achieved high residual turnover in the presence of Cmpd. 2 (Table 3b) and DKN (Table 3d). These results are also seen by significantly improved ki values of SEQ ID NO: 31 (Table 4) compared to the HPPD polypeptide (SEQ ID NO:1), and prior art mutant HPPD polypeptide (WO2014/043435) corresponding to SEQ ID NO:2 in this invention.
{Tabelle 4 listet 31 HPPD-Polypeptide und und deren Inhibitionskonstanten: „Evaluation of tolerance of mutate HPPD polypeptides against different HPPD inhibitor herbicides belonging to various chemical classes by the determiniation of the ki values“}
[0261] Starting from mutant HPPD polypetide corresponding to SEQ ID NO:8, further change of amino acid 330 (see SEQ ID NO: 12) shows further improved HPPD inhibitor tolerance (see Table 3)
[0262] Starting from mutant HPPD polypetide corresponding to SEQ ID NO:12, further change of amino acid position 264 (see SEQ ID NO: 18) show further significant improved HPPD inhibitor tolerance towards Cmpd. 1 and Cmpd. 2 (see Tables 3a, 3b).
[0263] Introducing all above mentioned exchanges in the four positions 264, 268, 270, 330 on top of mutant HPPD polypeptide corresponding to SEQ ID NO:10 leading to SEQ ID NO:19, the strongest tolerance of all depicted polypeptides in Table 3a with residual activity and turnover of 68% and 79% towards Cmpd. 1 was detected, demonstrating the importance of the combinatorial residue exchanges at the disclosed amino acid positions. Also the mutant HPPD polypeptide corresponding to SEQ ID NO:19 having improved ki values for Cmpd. 1 and Cmpd. 2 (Table 4) compared to SEQ ID NO:2.
[0264] For clarity, empty cells at the respective amino acid position in SEQ ID NO:2 to SEQ ID NO:108 are defined as identical to the amino acids corresponding to SEQ ID NO:1, highlighting only the exchanges in the mutant HPPD polypeptides. The mutant HPPD polypeptides represented here are examples by a way of illustration, not by a way of limitation. Data were obtained by measuring the initial reaction rates with increasing concentrations of Cmpd. 1 (2-chloro-3-(methylsulfanyl)-N-(1-methyl-1H-tetrazol-5-yl)-4-(trifluoromethyl)benzamide), Cmpd. 2 (2-methyl-N-(5-methyl-1,3,4-oxadiazol-2-yl)-3-(methylsulfonyl)-4-(trifluoromethyl)benzamide), and mesotrione (MST) according to Example 3. Generally, six different concentrations of HPP substrate (0-1350 μM) and four different concentrations of the respective inhibitor were applied (see FIG. 4). The inhibitor concentrations were adopted for the respective HPPD polypeptide/inhibitor pairs to generate dynamic data, i.e. variants with lower tolerance were analyzed in a range of lower inhibitor concentrations, and concentrations of up to 1200 μM were used for variants with maximized tolerance. GraphPad Prism (version 6.00 for Windows, GraphPad Software, La Jolla Calif. USA) were used for data analysis and fitting of kinetic constants applying constraints according to a competitive inhibition mode. Where obvious outliers occurred, or activities obtained at very high substrate concentrations didn’t obey the mathematics underlying the competitive inhibition mode, respective values were excluded from the fit.
[0265] Some results of systematical variants generated on the basis of the two already significantly improved mutated HPPD polypeptides SEQ ID NO:17 and SEQ ID NO:19 are outlined in Table 5.
{Tabelle 5 listet 63 systematisch erstellte Varianten und deren Herbizidtoleranz: „Effect of single residue exchanges in SEQ ID No: 17 and SEQ ID NO: 19 analysed according to Example 3 at low substrate concentration and 50 µM inhibitor Cmpd. 1.“}
[0266] SEQ ID NO:17 and SEQ ID NO:19 differ at two amino acid positions, and SEQ ID NO:17 shows a ˜20-fold, SEQ ID NO:19 a ˜30-fold increased ki towards Cmpd. 1 compared to mutant HPPD polypeptide corresponding to SEQ ID NO:2 (see Table 4).
[0267] The mutant HPPD polypeptide corresponding to SEQ ID NO:17 has residue exchanges at positions 268, 270, 335, 336, 337, 340, and 345 and exhibits 29% residual activity and the substrate turnover is reduced by only 46% in the presence of 50 μM Cmpd. 1. A reversion of residues at position 268, 270, 340, 345 to the respective wild-type residue (according to SEQ ID NO:1) is attended by a drop in tolerance to 19%, 24%, 17%, and 20% residual activity, respectively (Table 5; SEQ ID NO:71, SEQ ID NO:63, SEQ ID NO:81, SEQ ID NO:88, respectively) emphasizing the advantageous properties of these positions and mutations. Accordingly, a reversion of the residue Valine at position 345 in SEQ ID:19 to the respective wild-type residue Isoleucine is attended by a drop in residual activity from 67% to 57% (Table 5; SEQ ID NO:93).
[0268] On the other hand, the introduction of further single residue exchanges into SEQ ID:17 at position 204, 213, 264, 310, 315, 330, 331, 338, 339 or 344 resulted for every position in at least one variant with a further significantly increased residual activity of greater than 38%, e.g. A204M (SEQ ID NO:40), R213L (SEQ ID NO:44), M264K (SEQ ID NO:67), Q310K (SEQ ID NO:65), T315R (SEQ ID NO:45), D330V (SEQ ID NO:38), D331I (SEQ ID NO:49), F338V (SEQ ID NO:53), K339E (SEQ ID NO:55) and S344P (SEQ ID NO:58).
[0269] In summary, specific additional residue exchanges beyond mutations 335 (glutamic acid (E)=>proline (P)), 336 (glycine (G)=>aspartic acid (D)/histidine (H)), and 337 (asparagine (N)=>serine (S)) in the HPPD polypeptides provide improvements in the herbicide tolerance and demonstrates the additional importance of the disclosed positions 204, 213, 264, 268, 270, 310, 315, 330, 331, 338, 339, 340, 344, 345 conferring tolerance improvements to HPPD inhibitor herbicides (Table 5). Finally, the combination of these disclosed positions lead to ki improvements across the different HPPD inhibitor classes, as demonstrated in Table 4 (e.g. SEQ ID NO:99, 100, 105, and 107) against Cmpd. 1 with more than 100-fold, against Cmpd. 2 with up to 90-fold, and at the same time up to ˜7-fold improvements against mesotrione (MST) compared to wild-type HPPD polypeptide (SEQ ID NO:1) and prior art mutant HPPD polypeptide (WO2014/043435) corresponding to SEQ ID NO:2 in this invention.
[0270] For clarity, empty cells at the respective amino acid position in SEQ ID NO:2 to NO:108 are defined as identical to the amino acids corresponding to SEQ ID NO:1, highlighting only the exchanges in the mutated HPPD polypeptide. Residual activities and residual turnover were determined according to Example 3 by measuring kapp and total change in signal respectively in the presence and absence of Cmpd. 1 (2-chloro-3-(methylsulfanyl)-N-(1-methyl-1H-tetrazol-5-yl)-4-(trifluoromethyl)benzamide) (50 μM) at low substrate concentration. For each mutant HPPD polypeptide the kapp and total change in signal without HPPD inhibitor herbicides served for normalization of the kapp and total change in signal in the presence of the herbicide, and resulting %-values are summarized.